Arnar Pálsson. „Hvort er líklegra að veiran sem veldur COVID-19 verði hættulegri eða hættuminni fyrir menn vegna stökkbreytinga?“ Vísindavefurinn, 1. apríl 2020. Sótt 15. apríl 2020. http://visindavefur.is/svar.php?id=79103.
Breytingar á erfðaefni leiða sjaldan til stökka í gerð eða hæfni lífvera[1] og flestar stökkbreytingar sem finnst í stofnum eru hlutlausar.[2] Stökkbreytingar sem skemma gen og draga úr hæfni eru kallaðar neikvæðar en þær sem auka hæfni lífveru á einhvern hátt eru kallaðar jákvæðar. Jákvæð breyting á veiru getur haft neikvæð áhrif fyrir hýsil hennar. Hlutlausar breytingar eru þær sem hafa engin áhrif á hæfni. Vísindamenn telja slíkar breytingar hins vegar mjög gagnlegar því þær nýtast til að rekja sögu stofna, tegunda og veira.
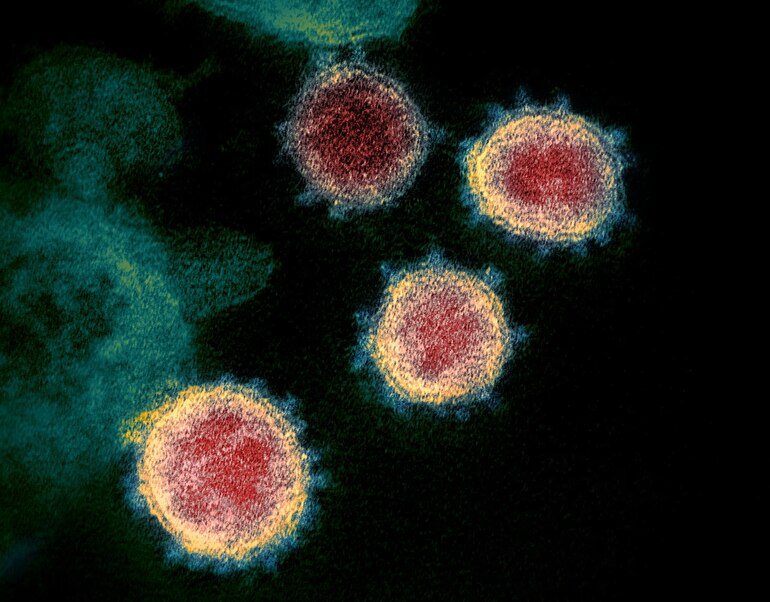
Lituð rafeindasmásjármynd af SARS-CoV-2-veirum.
Ef svo ólíklega vill til að þróunarlega jákvæð breyting verði í erfðaefni SARS-CoV-2 veirunnar sem veldur COVID-19, þá má spyrja hvort hún verði hættulegri við það? Umorða má spurninguna og spyrja: Hversu algengt er að veiran sem veldur COVID-19 stökkbreytist og geti við það orðið meira smitandi eða valdið alvarlegri sjúkdómum? Til þess að svara þessu þarf að skoða og skilja eftirfarandi:
- byggingu, lífsferil og erfðamengi veirunnar
- líkurnar á stökkbreytingum RNA-veira
- hversu líklegt það er að breyting sem er veirunni í hag valdi hættulegri sjúkdómi
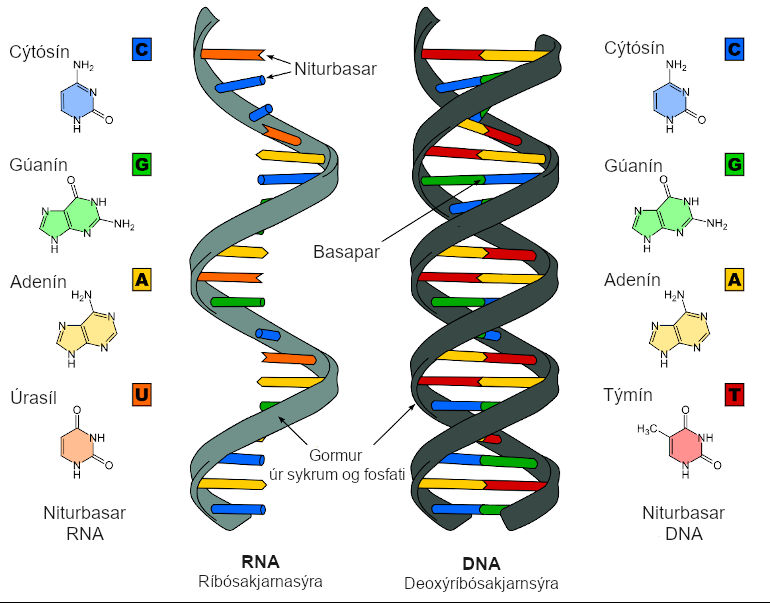
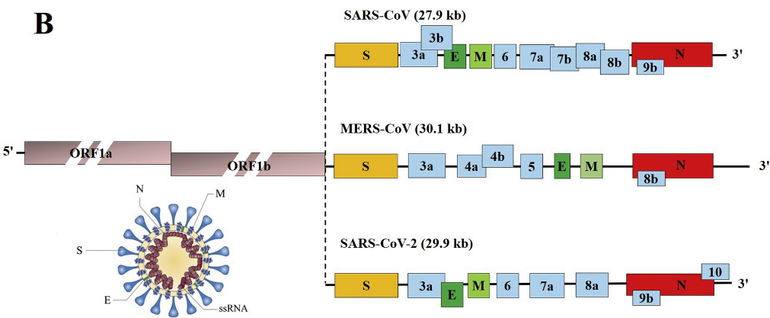
Um fyrsta atriðið er það að segja að veirur þurfa á lifandi hýsilfrumum að halda til að eftirmynda erfðaefni sitt og fjölga sér. COVID-19 er með einþátta 29.903 basapara RNA-streng.[3] Erfðamengi veirunnar skráir fyrir nokkrum fjölprótínum og minni prótínum. Eitt af þessum prótínum er mjög mikilvægt en það er svokallað bindiprótín (e. spike protein[4]). Það er á ytra byrði veirunnar og tengist viðtökum á yfirborði hýsilfruma, nokkurn veginn eins og lykill gengur að skrá. Veiran nýtir sér þannig tengingu bindiprótína við tiltekna viðtaka til innrásar í hýsilfrumuna. Síðan tekur veiran yfir kerfi frumunnar og fjölfaldar bæði erfðaefni sitt og nauðsynleg prótín og myndar að endingu fjölda nýrra veiruagna. Þær geta svo ýmist sýkt aðrar frumur sama einstaklings (þegar sýkingin magnast inni í manni) eða sloppið út í umhverfið og borist í aðra einstaklinga.
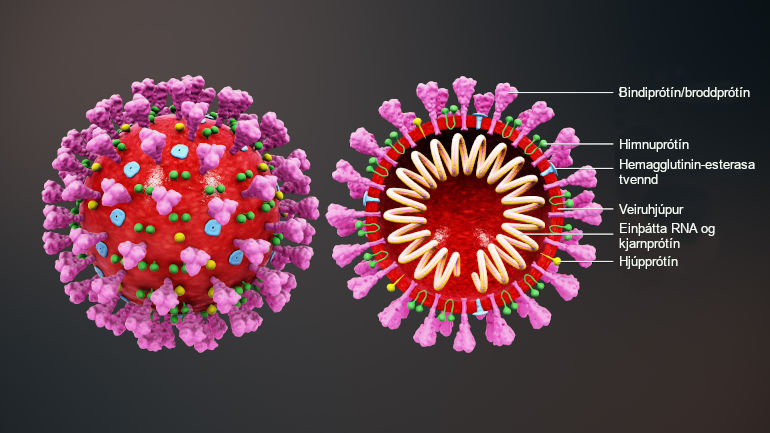
Einföld skýringarmynd af veirunni SARS-CoV-2. Bindiprótínin eru á ytra byrði veirunnar. Þau tengjast viðtökum hýsilfrumunnar og þannig kemst veiran inn í hana.
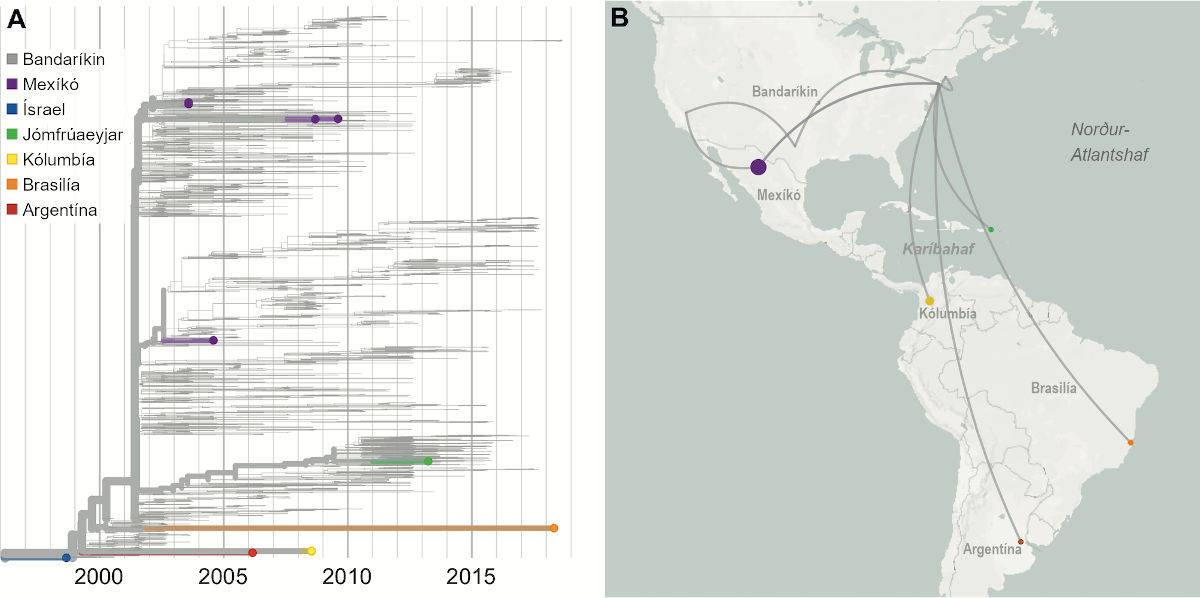
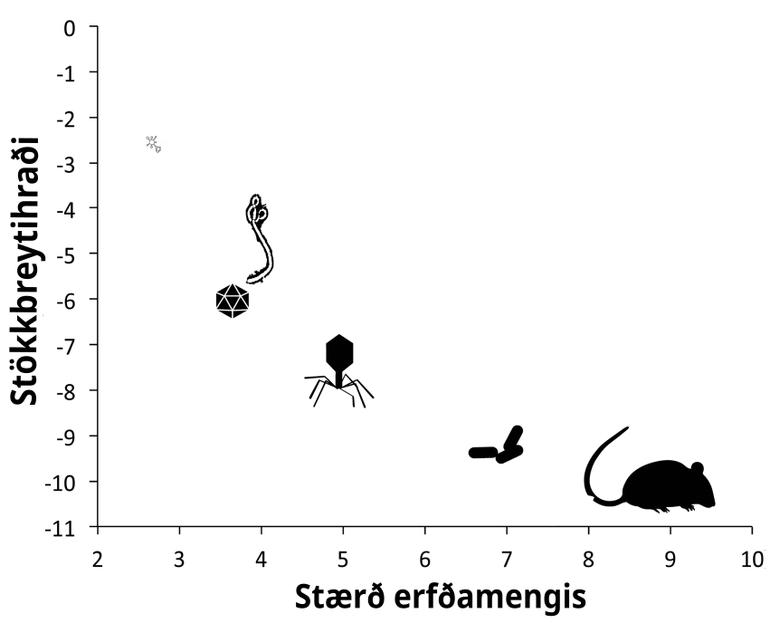
Um atriði tvö er rétt að hafa í huga að veirur, eins og lífverur almennt, mótast af kröftum þróunar, svo sem stökkbreytingum og náttúrulegu vali. Stökkbreytingar, sem réttara væri að kalla erfðafrávik, eru náttúrulegt fyrirbæri. Stökkbreytitíðni veira er há, sérstaklega miðað við heilkjörnunga og flóknar lífverur.[5] Vísindamenn um allan heim hafa raðgreint erfðamengi COVID-19-veirunnar úr meira en 1.500 smituðum einstaklingum á innan við tveimur mánuðum. Gögnin eru aðgengileg öðrum vísindamönnum í opnum gagnagrunnum.[6] Með erfðamengjum og ættartré veira úr smituðum einstaklingum er hægt að telja breytingarnar í hverri veirukynslóð. Í náskyldu kórónuveirunni SARS-CoV-1 er til dæmis tíðni breytinga á hvert basapar í hverri kynslóð 0,8 til 2,4 × 10-3. Við hvert smit þeirrar veiru urðu að meðaltali 16 breytingar í 30.000 basapara erfðamengi hennar. Gögn sameindalíffræðingsins Peter Thielen við John Hopkins-háskólann í Baltimore og samstarfsmanna hans, sýna að stökkbreytitíðni COVID-19 er ef til vill ívið lægri. Við smit frá kínverskum einstaklingum til bandarískra urðu 4 til 10 breytingar í erfðamengi veirunnar.[7] Stökkbreytitíðni er mæld sem fjöldi stökkbreytinga á kynslóð og því gefur auga leið að fjöldi smita hefur áhrif. Með auknum fjölda smita eykst hættan á að jákvæð breyting verði í erfðaefni veirunnar. Ef aðeins 1000 manns smitast þýðir það 1000 möguleika fyrir stökkbreytingar með jákvæð áhrif á hæfni veirunnar. Ef milljón manns smitast aukast líkurnar á breytingum sem eru veirunni í hag. Rétt er hins vegar að hafa í huga að líkurnar á slíkum jákvæðum breytingum eru fjarska litlar til að byrja með. Þá er komið að þriðja atriðinu. Ef svo illa vill til að jákvæð breyting verði á veirunni, getum við þá spáð fyrir um áhrif hennar eða hvort hún muni leiða til þess að veiran verði okkur hættulegri? Tvær afleiðingar væru alvarlegastar. Í fyrsta lagi að veiran breytist og valdi alvarlegri sjúkdómi (til dæmis með hærri dánartíðni) og í öðru lagi að hún smitist betur manna á milli. Mjög ólíklegt er að stökkbreyting komi upp sem valdi hvoru tveggja, meira smiti og hættulegri einkennum. Jákvæðar breytingar á erfðaefni veiru geta aukið hæfni í gegnum marga ólíka þætti lífsferils hennar. Til dæmis með því að bæta hæfileika hennar til að bindast frumum, til að sýkja fleiri gerðir fruma, til að framleiða fleiri agnir áður en einkenni koma fram hjá hýsli (auka smitmöguleika), lifa lengur í umhverfinu eða við lægra rakastig, valda tilþrifameiri hnerraköstum (sem auka dreifingu), valda heiftarlegri sýkingu, sleppa undan ónæmiskerfi hýsla og svo mætti lengi telja. Mjög erfitt er að spá fyrir um hvaða afleiðingar breytingar sem þessar (ef þær verða) hjá COVID-19 myndu þýða fyrir menn.

Fá dæmi eru þekkt um að stökkbreytingar eða erfðafrávik í veirum geri þær hættulegri. Stökkbreytingar á erfðaefni veiru sem leiðir til bráðs andláts hýsils veldur til dæmis minni hæfni hennar samanborið við óstökkbreytta veiru.
Fá dæmi eru þekkt um að veira stökkbreytist yfir í verra og svæsnara form. Best skilgreinda dæmið er zíkaveiran. Í henni varð stökkbreyting sem jók sækni veirunnar í taugavefi manna þannig að hún olli meiri skaða. Almennt er það veirum ekki í hag að drepa hýsla sína. Stökkbreyting í veiru sem leiðir til bráðs andláts hýsils veldur minni hæfni hennar samanborið við óstökkbreytta veiru. Ástæðan er einfaldlega sú að við það það dregur úr sýkigetu veirunnar. Því er talið að eftir að veirur aðlagist að endingu hýslum sínum valdi þær mildari sýkingum. Hinar fjórar kórónuveirurnar[8] sem sýkja menn valda til dæmis allar mildari sjúkdómi en COVID-19 og líklegt verður að teljast að COVID-19 þróist í svipaða átt og þær. Á hinn bóginn verða veirur að viðhalda sýkimætti og tryggja dreifingu sína (með hnerrum, hóstum og nefrennsli), þannig að það er veirum í hag að valda einkennum hjá sjúklingum sem tryggja dreifigetu veiranna. Veiran sem veldur COVID-19 þróast í því umhverfi sem hún býr í. Umhverfi hennar er maðurinn og samfélag okkar árið 2020. Viðbrögð samfélaga við veirunni eru hluti af því umhverfi. Einn vinkill að auki, er sá að ónæmiskerfi okkar er leið til að verjast sýkingum og muna eftir þeim. En sýklar geta einnig þróast sem viðbragð við auknu ónæmi meðal hýsla, eins og þekkt er hjá kórónuveiru sem nefnist OC43 (HCoV-OC43). Hún veldur aðeins venjulegu kvefi. Samantekt
- Stökkbreytingar eru í sjálfu sér ekki hættulegar.
- Stökkbreytingar sem gera veiruna svæsnari eru ólíklegar.
- Því lengur sem faraldurinn geisar því meiri líkur eru á breytingum sem eru veirunni í hag.
- Breyting sem er jákvæð fyrir veiruna leiðir ekki endilega til alvarlegri sjúkdóms.
- Fjórar aðrar kórónuveirur sýkja menn að staðaldri og leiða allar til mildari sjúkdóms.
- Að endingu er búist við að þróun veirunnar sem veldur COVID-19 leiði til mildara forms hennar.
Tilvísanir:
- ^ Arnar Pálsson. Geta lífverur þróast í stökkum vegna stökkbreytinga? Vísindavefurinn, 5. mars 2020. (Sótt 30.03.2020).
- ^ Arnar Pálsson. Getur kórónuveiran sem veldur COVID-19 stökkbreyst og orðið hættulegri?Vísindavefurinn, 23. mars 2020. (Sótt 30.03.2020).
- ^ Arnar Pálsson. Hvaðan kom COVID-19-veiran? Vísindavefurinn, 16. mars 2020. (Sótt 30.03.2020).
- ^ Önnur íslensk þýðing á orðinu er broddprótín.
- ^ Arnar Pálsson. Stökkbreytast veirur hraðar en flóknar lífverur? Vísindavefurinn, 23. mars 2020. (Sótt 30.03.2020).
- ^ GenBank er dæmi um gagnagrunn sem var settur upp til að vísindamenn gætu samnýtt gögn. Hann og aðrir sambærilegir grunnar eru mjög mikilvægir fyrir samþættingu upplýsinga, öllu samfélagi manna til hagsbóta. Hugmyndafræði opinna vísinda hefur verið mjög mikilvæg í baráttunni við COVID-19.
- ^ Sjá frétt Scientists say the coronavirus is not mutating quickly and might respond to a single vaccine á MSN endursögð úr The Washington Post.
- ^ Hér er ekki átt við veirurnar SARS-CoV-1 og MERS-CoV sem stukku yfir í menn og ollu afmörkuðum faraldri en var síðan að mestu útrýmt.
Heimildir og myndir:
- Grubaugh, N.D., o.fl. (2020). We shouldn’t worry when a virus mutates during disease outbreaks. Nature Microbiolgy 5, 529–530. (Sótt 30.03.2020).
- Ren, L., o.fl. (2015). Genetic drift of human coronavirus OC43 spike gene during adaptive evolution. Scientific Reports 5, 11451 (Sótt 30.03.2020).
- Yuan, L., o.fl. (2017). A single mutation in the prM protein of Zika virus contributes to fetal microcephaly. Science. 17.11.2017: 358 (6365), 933-936. (Sótt 30.03.2020)
- Kupferschmidt, K. (2020). ‘A completely new culture of doing research.’ Coronavirus outbreak changes how scientists communicate. Science, 26.02.2020. (Sótt 30.03.2020).
Myndir:
- Novel Coronavirus SARS-CoV-2 | This transmission electron mi… | Flickr. (Sótt 31.03.2020).
- 3D medical animation still shot showing coronavirus structure - Scientific Animations. (Sótt 31.03.2020).
- DNA Origami by Alex Bateman @alexbateman1 @sangerinstitute… | Flickr. (Sótt 31.03.2020). Myndin er birt undir leyfinu Creative Commons — Attribution 2.0 Generic — CC BY 2.0.