Measurement and Interpretation of the Microstructure of Fish Otoliths
Information from this page should be cited as: Campana, S.E. 1992. Measurement and interpretation of the microstructure of fish otoliths, p. 59-71. In D.K. Stevenson and S.E. Campana [ed.] Otolith microstructure examination and analysis. Can. Spec. Publ. Fish. Aquat. Sci. 117.
You can also download this chapter as a pdf file: Stevenson and Campana - Chapter 4
Introduction
A typical look at an otolith microstructure preparation reveals many ring-like structures, only a fraction of which are daily growth increments. While the interpretation of daily increments is more firmly based now than was the case even 6 years ago, an undeniable and substantial element of subjectivity yet remains. This point was underlined in a recent study, in which large numbers of known-age, mesocosm-reared herring (Clupea harengus) larvae of various ages were distributed for age estimation to experienced otolith readers representing 12 different countries (Campana and Moksness 1991). Larval ages and sampling frequencies were completely unknown to the otolith readers. On average, inter-sample age differences were accurately estimated in the study. However, there were significant (and occasionally large) differences among the readers, most of which could be ascribed to differences in training, technique and increment interpretation. An important conclusion of the study was that certain practices, both technical and interpretational, were superior to others, and that accuracy and precision, both among and within otolith readers, could be improved through reference to a standard protocol. This paper will attempt to provide some recommendations concerning otolith microstructure interpretation in order to meet, the above goal.
In the following sections, a number of guidelines will be offered as aids to the successful interpretation and quantification of otolith microstructure preparations. Most of the guidelines have a theoretical basis, but are allied with solid empirical support. Emphasis will be directed towards light microscopic images, since this is the medium most commonly used in microstructural examinations. For similar reasons, the discussion of quantification techniques will focus on daily increment counts and measurements. Other forms of measurement, such as isotope analyses, will not be discussed here. The reader is referred to Neilson (this volume) for additional information on sources of error associated with light microscope examination and interpretation of otolith increments.
Increment Counts
Currently accepted concepts of daily increment formation state that daily increments form as a result of an endogenous circadian rhythm (Mugiya et al. 1981; Campana and Neilson 1985). While environmental masking by fluctuating variables such as temperature and feeding may occur, such environmental cues tend to produce increments in addition to those produced as a result of the endogenous rhythm. Thus, the otolith microstructure can be expected to appear as a regularly-recurring daily increment sequence, occasionally overlain by or interspersed with subdaily increments of environmental origin. A useful analogy might be that of a regularly-recurring pattern of waves on the ocean (the endogenous circadian rhythm), overlain by waves and ripples resulting from passing boats (the environmental cues). The phase and amplitude of the waves due to boats would, of course, vary with their size and time of passing, resulting in boat-induced waves which could be either smaller or larger than the oceanic waves. In addition, the apparently-random phases and, wavelengths of the boat waves could amplify, negate, or intersperse with the oceanic waves, resulting in an overall wave pattern which may or may not appear regular. Subdaily increments form a similar pattern, in that they may be of variable intensities, widths and phases within a daily increment. However, the underlying daily increment pattern is usually smooth and regular. These observations, which are consistent with the underlying conceptual basis for increment formation, simplify the interpretation of previously-unexamined otoliths since microstructural growth patterns appear to be ubiquitous among all species (Campana and Neilson 1985; Jones 1986).
Selection of Counting Axis
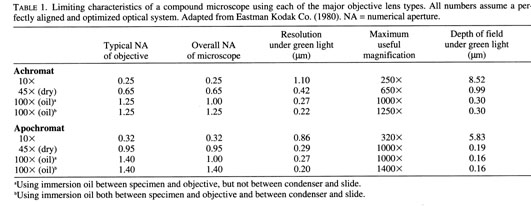
Selection of an appropriate counting path or axis is a mandatory step prior to further examination. Two criteria should be considered in the selection process: axis length and increment clarity. Axis length is a key factor, since not all otolith radii exhibit a complete increment sequence, particularly along the shorter axes (Fig.1). Sectors containing incomplete sequences can recognized by the presence of confluent daily increments and/or checks, and should be studiously avoided. While in theory it would be preferable to select the longest axis for examination, potentially-confusing increments will often be most prevalent along this axis. A practical compromise generally involves the shortest radius consistent with a complete increment sequence (Fig. 1). Selection of the counting path then be tempered by the second criterion, that of increment clarity. Variations in increment clarity along counting axis are the norm, and may be due either to inconsistencies in preparation or to uncontrolled factors associated with otolith growth. Since increment counts need not be carried out along a straight line, it is often helpful to shift the counting axis where required to avoid regions of ambiguity or poor clarity. Of course, increment continuity must be maintained at all times. When examination is to be made with a light microscope, counting paths are best first mapped out in the mind using magnifications that are lower than required for the subsequent increment counts.

Fig. 1. Selection of an increment counting path (solid line) in a polished juvenile cod lapillus. The path begins at the hatch check, proceeding distally to a check which serves as a landmark when the counting axis is shifted. While there are a number of possible counting paths, all would avoid an extended region of confluent increments (C) and a second region of unclear increments (U). Bar = 20 µm
Image Optimization
Microstructural features of most otoliths are minute; thus, subtle refinements in observational technique can prove to be of substantial value. Careful preparation of the sample, particularly through polishing, is generally most influential in enhancing visibility of an increment sequence (see Secor et al., this volume). However, the influence of proper microscopic technique, and more recently, image analysis systems, cannot be overstated. In what follows, emphasis will be directed towards optimization of light microscopic images, since this is the medium most commonly used in microstructural examinations. Optimization of SEM images is addressed in most SEM (scanning electron microscope) operating manuals, and will not be discussed here.
Microstructural examinations are best made with a compound microscope with the following features (at a minimum): binocular eyepieces, at least one of which can be focused; objective lenses with nominal magnifications of 16, 40 and 100X, at least one of which is designed for oil immersion; a moveable specimen stage; a substage condenser lens; an aperture diaphragm; and a variable-intensity illuminator with its own focusing/condensor lens. Use of an inferior microscope will, at best, introduce eyestrain in the observer, and at worst, introduce substantial error into the increment count. The most serious risk involved in the use of a microscope with inadequate or poorly-aligned optics is the failure to recognize the presence of narrow increments (<1 µm in width), such as would occur in temperate fishes with a pelagic larva stage (Campana et al. 1987; Jones and Brothers 1987) and subadult/adult fishes.
Image quality in general, and resolution in particular, can be influenced as much by microscopic technique as by hardware. For this reason, and given the sensitivity of many otolith interpretations to the quality of the image, some discussion of the factors influencing resolution are warranted. Resolution is defined here as the minimum distance between two structures consistent with the two structures remaining visually discernable. For a more complete discussion of microscopic principles and techniques, the reader is referred to one of the many excellent texts on the subject (eg. Eastman Kodak Co. 1980).
The objective lens is probably the single most influential factor in modifying resolution. Of the three major types, the achromat lens is the most popular and the least expensive. Such a lens provides partial correction for colour and spherical aberration, as well as a low but serviceable numerical aperture (NA). (The lens type, objective magnification, and NA are invariably etched into the body of the objective). Users of an achromat lens will often note that the image appears to improve when the aperture diaphragm is closed down, and when light of a single colour (e.g., green light) is used. The change in image quality can be attributed to the fact that the lens is not fully corrected for all wavelengths of light or for the entire field of view. The semi-apochromat lens, generally made of fluorite, is more completely corrected for aberrations, while the apochromat lens is almost fully corrected. The latter provides both the best image quality and resolution (Table 1), although all three types can be used for routine examination of otolith microstructure. All lens types can be purchased as flat-field objectives (e.g., planoachromat), which improves the image towards the edge of the field of view.
The numerical aperture (NA) of the objective ultimately controls both the magnification and the resolution that are obtained. Resolution (R) increases with NA as in:
where lambda is the wavelength of light that is used. Thus, the highest resolution is possible with objectives of the greatest NA (Table 1). Note, however, that the NA of the microscope is limited by the refractive index of all media between the condenser and the objective, as well as by the NA of the objective lens. The presence of air along the light path limits the effective NA to 1.0, no matter how large the NA of the objective being used. Therefore, an oil immersion objective must be used if an NA > 1.0 is to be achieved. What is not as widely known is that immersion oil must also be used between the condenser lens and the bottom of the microscope slide for an overall NA > 1.0 to be reached. Few microscope users (in the field of otolith microstructure) appear to be aware of this constraint, making the useful resolution limit of a perfectly-aligned light microscope close to 0.27 pm (Table 1). The use of a blue filter over the light source can improve this limit by 15-20%.
While resolution has a well-defined limit in light microscopy, magnification can be increased almost endlessly. Thus, it is fairly easy to set up a microscope with a 100X objective lens, 25X eyepieces and a 2X body tube to yield a magnification of 5000X. However, most of the magnification is "empty"; that is, the image is large, but reveals no extra detail beyond that visible at around 1250X. The maximum useful magnification for most microscopes is 1000-1250X (Table 1).
Aside from the objective, an optimized source of illumination will have the greatest influence on image quality. Kohler illumination is the most common means of optimizing a light source, and will result in bright and even illumination over the entire sample, with good depth of field and resolution. The steps involved in adjusting for Kohler illumination are discussed elsewhere (Eastman Kodak Co. 1980), but revolve around centering the light source in the image, focusing the light on the plane of the stage (by focusing the condenser), and adjusting the aperture diaphragm. A properly focused condenser will generally be near the top of its travel range. Once in place for a given microscope, few extra adjustments are needed as the specimen or magnification are changed. While it is often used (incorrectly) to compensate for changes in illumination, a properly adjusted aperture diaphragm will balance contrast, depth of field and resolution.
Once the optics of the microscope have been aligned and optimized, the otolith reader must select the magnification which will be used to examine the sample. The intent should be to balance the apparent clarity of the increment sequence with ease of counting. Daily increments invariably appear most distinct at lower magnifications, in part because visual artifacts and subdaily increments (and any narrow daily increments) are less prominent. Aside from the reduced resolution of adjacent increments, it is intrinsically difficult to count sequences of growth increments at low magnification; the human eye tends to wander involuntarily when large numbers of structures with similar appearances are visible in the same field of view. At the other extreme, high-magnification examination of broad increments can be very confusing; the internal structure of both the discontinuous and incremental zones can be surprisingly complex. As a general rule of thumb, a magnification of 400X is often appropriate for the examination of rapidly-growing otoliths, while 1000-1250X will be mandatory where increments are less than 1-2 µm in width. In both instances, a useful endpoint is a field of view where about 20 increments are visible at one time. Fortuitously, the use of immersion oil with high magnification objectives (those above 40X) tends to smooth out the image by obscuring surface imperfections in the sample.
While not necessarily useful in other applications of microscopy, frequent focal adjustments during the scanning and/or counting of growth increments are almost mandatory. Focal adjustments not only simplify the differentiation of daily and subdaily increments, but they compensate for intrinsic variations in the focal plane of the increments themselves. Such variations, whether due to alignment errors during mounting or to nonplanar otolith growth, account for the difficulties and/or inaccuracies that many workers experience in counting daily increments from photographs. The photographic depth of field is also very small (Table 1). For this reason, direct microscopic examination is generally preferred over photomicrography for counting increments.
Image Analysis Systems
An image analysis system can be a powerful tool to those working with otolith preparations. Thus, a brief description of the capabilities and applications of image analysis is warranted. Image analysis is a generic term used to refer to the digitization and manipulation of visual images. In its simplest form, an image analysis system can store a picture in memory and reproduce it, unaltered, upon command. In practise however, images entered into an image analysis system are enhanced and/or quantified before re-display; therein lies their advantage over visual examination. The end product is an image (or data) which can be more easily interpreted than the original. Image analysis systems should not be confused with simple video-microscope display units. While the latter have been used to advantage in studies requiring precise otolith measurements (i.e., Methot and Kramer 1979; Bolz and Lough 1983), such units are capable of neither image enhancement nor image manipulation.
Recent technological advances have brought microcomputer-based image analysis systems within the financial grasp of an individual researcher. Standard systems now consist of a video camera, a digitizer board mounted within a microcomputer, and a monitor (for an example, see Campana 1987). The video camera would be mounted on a microscope for otolith-based research. Most of the digitizer boards available today are "framegrabbers", capable of digitizing and storing 30 images per second. Thus, real-time viewing and image manipulation is not only possible, but the norm.
The basis of operation for all image analysis systems is the conversion of an image into an array of numbers - in other words, image digitization. Each position in the array represents a pixel (grid square) in the image, and each numerical value represents a gray level (measure of light intensity) for that pixel. Thereafter, anything that can be done to an array of numbers can be done to an image. For example, image contrast can be doubled by doubling each pixel's gray level. Since the results of an array manipulation can be seen immediately on the monitor, image manipulation can be as interactive or as automated as desired.
In the context of microscopic observations, image analysis systems provide three major advantages: image enhancement, manipulation, and quantification. Image enhancement is one of the most important and widely-used features. Simple procedures allow the operator to subtract an image background from the entire image, average several noisy images, or use high or low-frequency filters to add or remove detail. Gray level expansion, whereby the gray levels in a poorly-contrasted image are spread out over all 128 (or more) levels, can bring out detail that is totally invisible to the unaided eye (Fig. 2). All of these enhancement procedures are effective because of the limited capability of the human eye - differentiation of 128 gray levels is well beyond our visual capacities.
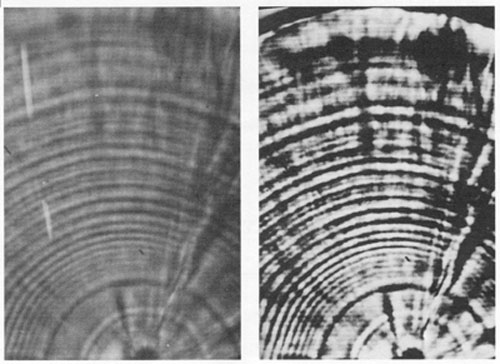
FIG. 2. Grey level expansion (right), with an image analysis system, of a poorly-contrasted light-microscopic view (left) of an otolith growth sequence, as photographed directly off of the video monitor. Both photographs were taken, developed and printed under identical conditions. Preparation of the enhanced video image took approximately 2 seconds. While grey level expansion is an effective means of enhancing contrast, other image analysis procedures can be used to further sharpen or filter an image.
Increased ease of visual interpretation is the primary advantage of an image analysis system in counting growth increments. When measurements are being made, a variety of other features become evident. Foremost of these is ease of measurement. Not only is a video monitor target easier to position than an ocular micrometer, but distances approaching the theoretical resolution limit of light microscopy (0.20 µm) can be measured. Of course, all measurements can also be stored directly in computer memory, eliminating the error potential of handwritten transcription. These and many other applications are detailed elsewhere (Gonzalez and Wintz 1977; Hall 1979; Ballard and Brown 1982; Campana 1987).
Despite the undeniable benefits of image analysis systems to otolith microstructure examination, it is important to recognize their limitations as well. Use of such systems does not improve resolution; while visual contrast can be enhanced considerably, the resolution limit of light microscopy is inviolable. Secondly, the automatic-count capabilities of many systems are not yet appropriate for studies of otolith microstructure. And finally, image storage presently requires too much memory (256K) for the creation of large image archives. Since image analysis technology is now progressing rapidly, these latter two constraints may well disappear in the near future.
Increment Interpretation
In a typical otolith preparation, numerous ring-like structures are evident, only a fraction of which are daily growth increments. With familiarity, the distinction between "real" daily increments and most other features is a routine procedure. However, unpractised otolith readers can introduce enormous errors into an increment count, while even practised workers can differ (sometimes substantially) in their interpretation of a given increment sequence. This element of subjectivity is one of the most significant sources of error in otolith microstructure examination, and largely explains the current absence of automated counting instruments. It also explains why validation (discussed in Geffen, this volume) is more important as a check on the interpretive skill of the worker, than as a check on the true frequency of increment formation. In this section, a number of guidelines will be offered as an aid to more informed interpretation of microstructural features. Not surprisingly, ease of interpretation improves with experience and the degree of sample preparation.
The three light-microscopic features most commonly confused with daily growth increments are, in order of importance: subdaily increments, visual artifacts, and checks. All except visual artifacts serve to confound SEM interpretations as well. Criteria for the differentiation of the three features are largely based upon visual appearance; in particular, contrast and relative width (Fig. 3). However, location on the otolith may also provide clues as to the identity of a particular structure. For instance, daily increment widths are often narrow in the region encircling the hatch check, while broad, clearly-defined increments characterize growth during the juvenile stage. As age increases beyond this point, increments tend to narrow, and may become vanishingly small, or even intermittent in their formation. Knowledge of this general growth pattern is often helpful in interpreting an increment sequence (Fig. 4).
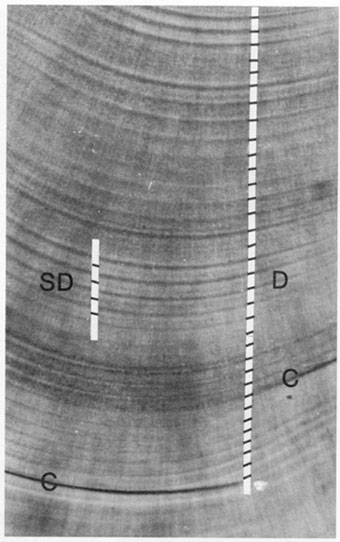
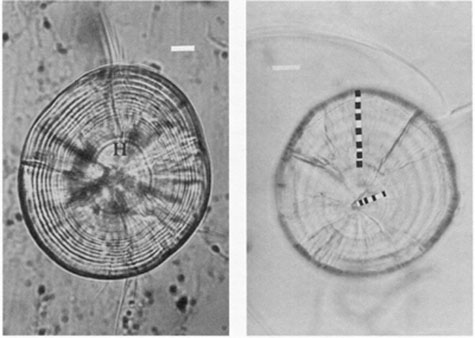
FIG. 4. Daily increments can often be differentiated from similar-appearing structures by their location on the otolith and by species-specific characteristics. (left) Daily increments encircling the hatch check (H) of a polished haddock sagitta are narrow and weakly expressed for the first 10-20 d after hatch, but broaden as the postlarval and juvenile stage is entered. Many of the perinuclear daily increments visible under greater magnification with the light microscope are not evident in this photograph, which shows only 24 of the total of 39 increments from a 8.35 mm larva. Bar = 10 µm. (right) In contrast to the haddock sagitta, the lapillus from an 11-d old walleye larva has broad daily increments almost from the date of hatch. Broad increments such as these are also characteristic of many tropical fish otoliths. In such cases, low magnifications (300-500X) may be most appropriate for the examination. Bar = 20 µm.
Daily and subdaily increments are morphologically similar, making differentiation of these two types of structures difficult. Criteria for their differentiation take advantage of the differing factors behind their formation. Daily increments form at a constant frequency, and due to the conservative nature of otolith growth, often appear as a regular sequence with smooth transitions in both increment width and increment contrast. Subdaily increments on the other hand, may form at any date or time of day, rendering their widths less regular. The visual prominence of subdaily increments is usually less than that of adjacent daily increments (Fig. 3), but will vary with the strength of the masking agent responsible for their formation. In practice, it is often best to locate a region of unequivocal daily increments along the intended counting path, and then proceed outwards (or inwards) from that point, using the regularity/continuity criteria in interpretation. Where adjustment of the microscope focus appears to "split" increments, the broader of the two patterns can often be assumed to be daily. Indeed, in cases where otolith growth has been rapid and subdaily increments are numerous, a slightly out-of-focus examination may aid in eliminating subdaily increments from the field of view (Fig. 5). This approach is not appropriate where increments are narrow, such as around the hatch check, since aggregates of daily increments may then become evident (Fig. 6). Where ambiguity between daily and subdaily increments persists, the best tactic is to avoid that region of the otolith. Counting paths need not be linear, and interpretive ease of a given sequence often varies among the potential counting axes.
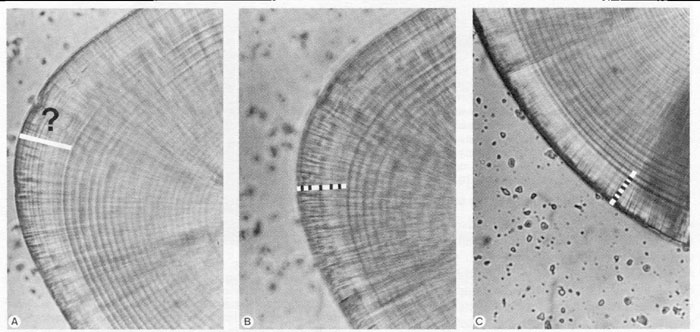
FIG. 5. Tactics for dealing with "split" or subdaily increments. (A) A properly-focused view of the edge of a polished herring sagitta. Whether due to preparation difficulties or to the presence of subdaily increments, an unambiguous sequence of daily increments from nucleus to edge suddenly shifts to "daily" increments (11 or more) of much narrower width. The sudden shift in increment widths, along with the apparent splitting of the most medial increment of the marked zone, indicates that the zone should be interpreted carefully. (B) The same view as in (A), intentionally made out of focus. The periodicity of the broad, underlying pattern in the suspicious zone is similar to that of the unambiguous daily increment sequence, suggesting that the zone actually represents 4 or 5 daily increments. (C) A different region of the edge of the same otolith visible in (A) and (B). This region of the otolith confirms the increment interpretation derived from the out-of-focus examination in (B).
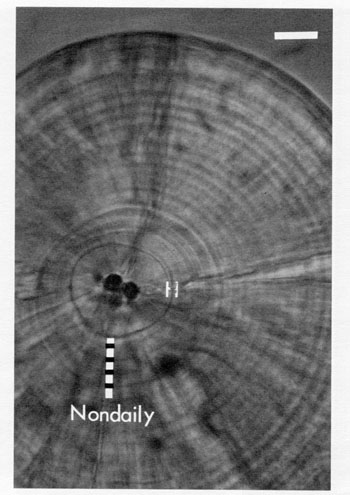
FIG. 6. Aggregates of daily increments may sometimes look more convincing than do the actual daily increments. The 4 false increments marked on this photograph of a known-age herring larva sagitta actually correspond to a period of about 20 d. Some of the true daily increments are visible to the upper right of the hatch check (H). Careful and frequent adjustments of the microscope focus are required to correctly interpret this type of increment sequence. Familiarity with the overall growth pattern of herring otoliths would also prove useful here. Bar = 10 µm.
Subdaily increments tend to be most prevalent in two situations. The first is in regions in which the otolith has grown rapidly, resulting in very broad daily increments. The broader the incremental zone of a given increment, the greater the potential for subdaily increments to have formed, and more importantly, the easier it is to see them. This is one reason why the microstructure of a juvenile fish sagitta can be difficult to interpret - the increments tend to be very broad (Fig. 7). Since the lapillus has a lower and more spatially uniform specific growth rate, increments are narrower, and as a result, better defined (Fig. 7) (see Secor et al., this volume, for further detail). Of course, this inter-otolith growth difference can be counterproductive in lapilli of old fish, since increments there can be so narrow so as to be difficult to resolve.
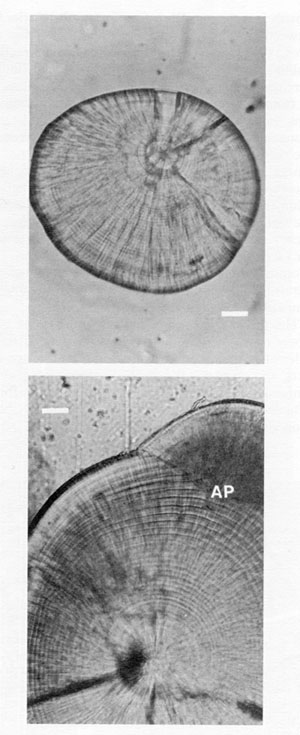
FIG. 7. A comparison of the microstructure of lapilli and sagittae from the same, 20 mm cod. Both otoliths have been polished, and reproduced at the same scale. Bar = 20 µm. The growth sequence in the lapillus (top) has well-defined and spatially-uniform increments, although the latter would become increasingly narrow and difficult to interpret in older juveniles. In contrast, the daily increments in the sagitta (bottom) are narrower than those of the lapillus for the first 5-15 d after hatch (not visible at this magnification), but become increasingly broad with age. Increments towards the edge of the sagitta are more than 3 times as broad as those at equivalent ages in the lapillus; the sagitta also shows evidence of splitting and/or subdaily increments in the outermost 15 d. Daily increments are broader yet, but indistinct, around the newly formed accessory primordium (AP) at upper right.
The second situation promoting visibility of subdaily increments is an artifact of preparation difficulties. For various reasons, overgrinding can make subdaily increments appear more prominent than daily increments. Indeed, the former can appear both regular and well-defined in overground preparations, making this a particularly dangerous sequence to interpret (Fig. 8). Proper recognition of overground regions can minimize counting inaccuracies due to this effect.

FIG. 8. Overgrinding can make subdaily increments appear more prominent than they would be otherwise. Subdaily increments are prominent, and daily increments indistinct, along two marked growth axes of an overground starry flounder (Platichthys stellatus) sagitta. Daily increments have been indicated in black; the daily pattern is also more apparent in the better-prepared section to the lower right.
Visual artifacts take several forms, some of which may mimic daily increments. Refraction of light through and around the curved edge of the otolith can distort the width and/or number of increments visible, making accurate interpretation difficult. However, edge effects usually compromise the appearance of only a few increments, thus allowing interpolation if necessary. Artifacts resulting in increment "reflections" are most visible just outside the perimeter of the otolith, but are also associated with checks and prominent increments. Differentiation is on the basis of the appearance of the adjacent structures; artifacts appear as exact reflections, sometimes in multiple copies, of the nearest increment, but are usually more sharply defined than the reflected structure (Fig. 9). Focal adjustments can serve to minimize the number of visual artifacts, but since they may also influence the interpretation of true daily increments, should not be used as the sole defining criterion.
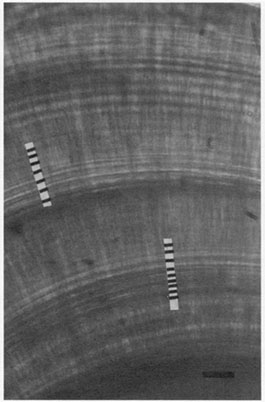
FIG. 9. While the fine lines between the marked daily increments in this photograph of a starry flounder sagitta were originally termed subdaily increments (Campana and Neilson 1982), their appearance under a light microscope and their absence under a scanning electron microscope indicates that they are actually visual artifacts associated with prominent increments and/or checks. Bar = 10 µm.
Checks have never been adequately defined, due in part to the variety of agents attributed to their formation (Pannella 1980; Campana 1983, 1984; Gauldie 1988). Where they appear as particularly prominent increments, perhaps in response to short-term stress (Campana 1983), interpretation is not a problem. Checks associated with the lunar cycle (Campana 1984) or interrupted otolith growth (Pannella 1980) are easily confused with cracks or fissures, and may appear to reside on a different focal plane than that of the surrounding increments (Fig. 3). Such checks are not daily increments, and should not be counted as such, although they may overlay true increments. Regions of interrupted otolith growth, perhaps characterized by confluent or rapidly-narrowing increments, should be avoided during increment counts. If a complete sequence around the problem area cannot be found, the otolith must be discarded; after all, there is no way to assess the duration of the otolith growth interruption. Note, however, that checks that overlay an apparently normal increment sequence seldom signify the presence of interrupted growth (e.g., Fig. 3).
While the presence of accessory primordia can complicate measurements of increment width (as discussed in a later section), they should have little effect upon increment counts. Increments are almost invariably contiguous across the growth zones corresponding to different points of nucleation (e.g., Fig. 7B, 11).
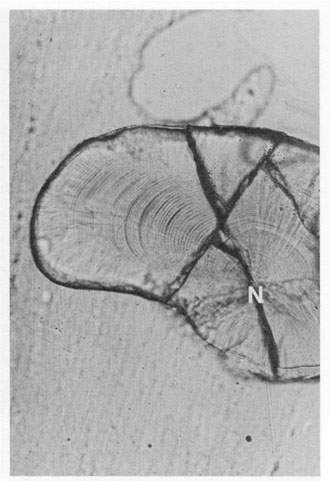
FIG. 10. Curvilinear growth axis in the lapillus of a juvenile flyingfish, Paraexocoetus brachypterus. There is no single straight line which can be drawn from the nucleus (N) to the otolith edge which will intersect a complete sequence of growth increments at right angles. However, all standard growth backcalculation procedures assume a linear backcalculation trajectory.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Optimization of Counts
The practice of providing a single best increment count for a given otolith can be unexpectedly difficult. Most scientists are capable of counting up to 100, or even beyond. However, increment counts are invariably complicated by interpretive difficulties, variations in preparation quality along the counting path, nonlinear counting paths, and the limitations of the human eye. In this section, we provide some empirically derived suggestions for minimizing these problems.
The human eye tends to wander involuntarily when counting extended sequences of tightly-packed, repeated structures such as growth increments. Distinct otolith features such as checks, scratches, or prominent increments should thus be used to advantage as stopping points or landmarks for the eye. Counting bursts of 10-15 increments between landmarks are ideal, since they allow the examiner to scan the region ahead for interpretive difficulties without losing track of the last counted increment. Such landmarks are also useful when lateral shifts of the counting path are to be made. Note also that constant adjustment of the focus while scanning not only simplifies the differentiation of daily and subdaily increments, but compensates for variations in the focal plane of the increments.
Preparation quality is seldom uniform along the counting path. In most cases, a complete count will be possible. However, there may be instances where cracks, overgrinding or undergrinding occlude short sections of the increment sequence. Since daily increment widths tend to be autocorrelated, interpolation may be justified if the interpolated increment number is very small relative to the overall count. Age estimates based on 5% interpolation are probably acceptable, although the exact percentage is arbitrary. Percentages as high as 20% have been reported (Methot 1983), but are not recommended. Interpolation is often appropriate at the otolith edge; however, it is not appropriate where increment widths are changing (i.e., around the nucleus).
A minimum of two complete counts (and preferably more) should be obtained for each otolith. Counting efforts will normally be minor compared to differences in interpretation. Since interpretation may differ with the point of origin for the counting path, one count should originate at the otolith periphery, while the other should begin at the hatch check (or designated first increment). There is no strict convention concerning the inclusion of the hatch check and the otolith periphery in the increment count; however, the method that is eventually adopted should always be reported. Calculation of a single "best" increment count for a given otolith is discussed in a later chapter (Campana and Jones, this volume).
Increment Measurements
Accurate measurements of daily increment widths are intrinsically more difficult than simple counts of the same increments. In addition, interpretation of the measurements in terms of otolith or fish growth is not necessarily straightforward (see Campana and Jones, this volume). In what follows, guidelines for the selection of a measurement axis and the collection of accurate measurement data will be presented. Details of image optimization and interpretation are similar to those discussed earlier, and will not be addressed further.
Selection of Measurement Axis
Selection of a suitable measurement axis requires the same axis length and increment clarity criteria as those associated with increment counts. However, there are two additional constraints put upon the selection procedure: the measurement path must be linear, and otolith growth should be roughly symmetrical. These constraints are based upon the eventual application of increment width measurements to calculations of otolith or fish growth. To be interpreted, increment widths must be put into the context of overall otolith size and/or growth rate. Yet virtually all otoliths have eccentrically positioned nuclei, implying that the width of a given increment can be expected to change with the length and orientation of the otolith radius under examination. Thus, a single, linear axis must be used for all increment width measurements within a given otolith. When the increments of more than one otolith are to be measured, the orientation of the measurement axis should be standardized to minimize among-fish variation. Note that the above requirements are far more constraining than those associated with increment counts, since regions of difficult interpretation or poor preparation quality cannot be avoided through lateral shifts of the field of view. There is also an additional complication. In instances where the axis of otolith growth is curvilinear, it is impossible to measure maximum increment width (parallel to the growth axis) while maintaining linearity between the nucleus and otolith edge (Fig. 10). Since oblique measurements of increment width are meaningless, growth backcalculations of such an otolith would have to be based upon the length of the curvilinear growth axis. Although theoretically possible, curvilinear growth axes have seldom been measured, presumably due to the difficulty of defining a curved line which intersects all growth increments at right angles.
The second constraint, that of growth symmetry, is associated with measurement axes that pass through regions where the axis or rate of growth has shifted. For example, the width of a given increment may change substantially with proximity to an accessory primordium (Fig. 11). Accessory primordia are seldom observed in lapilli, unlike the situation in sagittae, making the former a more suitable choice for increment width measurements. However, shifts in growth axes may occur in the absence of accessory primordia. For instance, many otoliths are nearly circular in larval fishes, but become increasingly asymmetrical through the juvenile stage. Thus, what may be a perfectly suitable measurement axis in a larval fish may be completely unsuitable in a juvenile. Such an effect can be controlled by ensuring that the measurement axis used for the juveniles is the same as that measured for the larvae. Since the two axes will not necessarily correspond with the longest axis of the otolith (at either life history stage), some care must be taken to ensure that the appropriate axes are used. The ubiquity of asymmetric otoliths also indicates that increment widths, by themselves (without an accompanying fish-otolith relationship) are poor indicators of growth rate.
Optimization of Measurements
Increment widths may be measured from SEM micrographs, light micrographs, digitized images, video displayed images, or using a light microscope with an ocular micrometer. Irrespective of the method used, individual increment measurements are predicated upon orientation parallel to the axis of growth at that increment, not just parallel to the overall growth axis. All techniques suffer from potential sources of bias or difficulty in preparation, although some problems are more acute than others. The selection of measurement technique should therefore be based on the requirements for accuracy and precision, as well as access to specialized equipment.
The exact procedure by which individual increments are measured is probably less important than is the consistency in procedure across increments. That is, there is little reason to believe that the measurement of an increment from the medial side of one discontinuous zone to the medial side of the next discontinuous zone will be any more (or less) accurate than, say, from the center of one incremental zone to the center of the next. However, whichever protocol is applied, it is very important that it be applied consistently to all of the increments under study.
The major source of bias in the measurement of increment width is the effect of the focal plane upon the image magnification and the resulting increment width. In light microscopy, adjustment of the focal plane is critical to obtaining an undistorted image of the increment sequence. Yet focal adjustments also induce shifts in the apparent width and position of each increment. Accuracy is maximized when the increment being measured is in optimum focus; since the optimum focal plane for one increment is not necessarily the same as that for the adjacent increment, compensations must be made for the consequent shift in increment position whenever the focus is adjusted. However, it is more difficult to compensate for the accompanying shifts in magnification. This problem applies as much to light micrographs as to visual microscopy. The best solution reported to date is to minimize or avoid the problem; the otolith should be mounted so that the incremental plane is as close to horizontal as possible. Small shifts in focus are unlikely to result in major changes in apparent increment width. However, the apparent lateral shift in increment position can be more substantial, and can result in significant measurement error if the new measurement start point is not used after refocusing.
Light-microscopic increment width measurements are best measured with an image analysis system (Campana 1987) or video-microscope system (Methot and Kramer 1979; Bolz and Lough 1983). While the former provides the added advantage of image enhancement capabilities, both provide the operator with a target on a large video screen, as well as the flexibility to make continual focal plane adjustments. Such measurements are much more precise than those obtained with an ocular micrometer, and reduce the potential for error by transmitting the data directly to a computer file. Video measurements are also much more rapidly obtained than those from an ocular micrometer or photographs.
The most accurate increment width measurements are derived from SEM micrographs. Such measurements are not subject to the refractive effects that can shift or distort an image under a light microscope. Accuracy and precision is then limited only by the clarity of the photograph and the means by which the increments are digitized. Aside from accessibility, the major constraint of SEM measurements is sample etching. Etching of a full increment sequence can be difficult to achieve (Blacker 1975; Campana and Neilson 1985). For this reason, SEM measurements are more appropriate for discrete regions of the otolith rather than complete radii.